
英语原文共 6 页,剩余内容已隐藏,支付完成后下载完整资料
体内早期基因表达即时成像揭示哺乳动物大脑中皮层特异性的记忆印迹
摘要:对于大脑皮层中长期记忆印迹形成的动态过程,人们知之甚少。这些过程的观测需要长时间检测大量的任务诱导的神经元。在此,我们提出了一种基于双光子成像的系统,通过EGR1基因启动子控制表达的可定量观测的EGFP蛋白来追踪数以千计的事件相关神经元的活动。我们开发了一种识别算法在不同皮层区域中检测GFP阳性的神经元,因此,可在2个月里对每一区域4000个神经元进行重复的追踪。结果分析显示,在稀疏的第二层中的神经元中具有环境特异性的反应。这种环境诱发的反应在数天的训练中逐渐增强,并且能够维持一个月。其形成的印迹可被训练环境特异性的激活,且和行为上的反应线性相关。对特定环境响应的神经元群很大程度上是分离的,表明在第二皮层中,记忆相关的印迹是稀疏编码的。
在哺乳动物大脑中,人们对皮层区域中的记忆印迹了解较少。相对于被认为与陈述性记忆短期存储有关的侧颞叶(medial temporary lobe),特别是海马(hippocampus)[1,2],新皮质(neocortex)被认为是用来存储长期记忆[3-6]。然而,由于皮层中神经元连接复杂、数量巨大,在细胞水平上揭示长期记忆存储的位置和动态过程相关的知识非常少。
体内神经元放电的电生理记录将神经元活动与动物行为联系起来,彻底改变了神经生物学。然而,电极记录的神经元数量少是一种限制,因为信息的编码和破译需要监测一群神经元形成细胞群[7,8]。为了记录皮层中更大数量的神经元活动,研究人员使用了不同的方法:增加电极数量[7,9-11],使用钙指示剂成像[12-15],即使早期基因(IEG)作为报告基因[16-18]。在外部刺激下,IEGs的表达与平均神经元活动相关[19-20],表明标记的神经元与行为有关[1,21-25]。体内IEGs成像研究揭示了视觉皮层和其他皮层中的编码,反映了单个神经元的电活动[16,17]。在IEGs中,早期生长应答蛋白1(EGR1,也被称为zif268)的表达与高频率的刺激和学习过程中长期可塑性的诱导有关[26,27]。为了监测行为任务中皮层环路的神经元活动,我们使用了一种EGR1表达报告基因的小鼠系,在这一小鼠系中,Egr1基因的启动子能够调控EGFP蛋白表达。通过对小鼠皮层细胞中EGFP信号的变化定量检测,我们设计了离线的(offline)记录策略来监测任务相关神经元的活动。在不同任务中激活的神经元集群模式在整个皮层中可视。此外,通过计算机识别重建,我们能够追踪来自多个皮层区域的与活动相关的细胞EGFP信号2个月,以揭示皮层回路中与记忆相关的变化。
意义
这一研究展示了哺乳动物大脑中,在复杂的行为中,感觉信息是如何在表示和存储在皮层回路之中。使用一种新建立的细胞自动检测算法,我们追踪了2个月来每只活老鼠超过20,000个神经元的即时早期基因的表达量,揭示了局部皮层回路中每个神经元的定量信号变化。一个自然的行为任务诱导皮层Ⅱ中稀疏的、任务特异性的神经元激活。这种逐渐形成的稀疏的神经元激活在多个皮层区域中被观测到,我们的研究结果支持皮质回路中长期记忆存储巩固的概念,并展示了自然事件在哺乳动物大脑中的稀疏编码。
结果
我们首先校准了体内外神经元兴奋诱导的EGR1-EGFP表达量。转基因小鼠体内EGFP蛋白和mRNA水平反映了Egr1基因的表达量(Fig S1)。尽管EGFP蛋白被认为是稳定的,但在培养的皮层神经元中,使用突出传递化学阻滞剂(MK801,CNQX或钠离子通道阻滞剂河豚毒素(TTX))后一小时内EGFP信号出现了明显的衰减(Fig S2A)。相反,使用二羟基苯甘氨酸(DHPG)增强神经元的激活在之后的一个小时内引起EGFP信号增强。因此,EGFP信号反映了培养神经元的激活。为了进一步了解活动依赖的EGFP表达的特征,我们定量测量了感染了光敏感通道蛋白2(ChR2)的皮层神经元对频率特异性刺激应答诱导的EGFP表达。在刺激一小时后测量显示,EGFP的表达量与光频率具有很大的正相关性(Fig S2B)。由于对表达ChR2的神经元刺激后,会诱导其以光刺激相同的频率放电[28,29],这种频率依赖的EGFP表达显示,神经元中EGFP水平记录前一小时刺激的峰值强度相关。
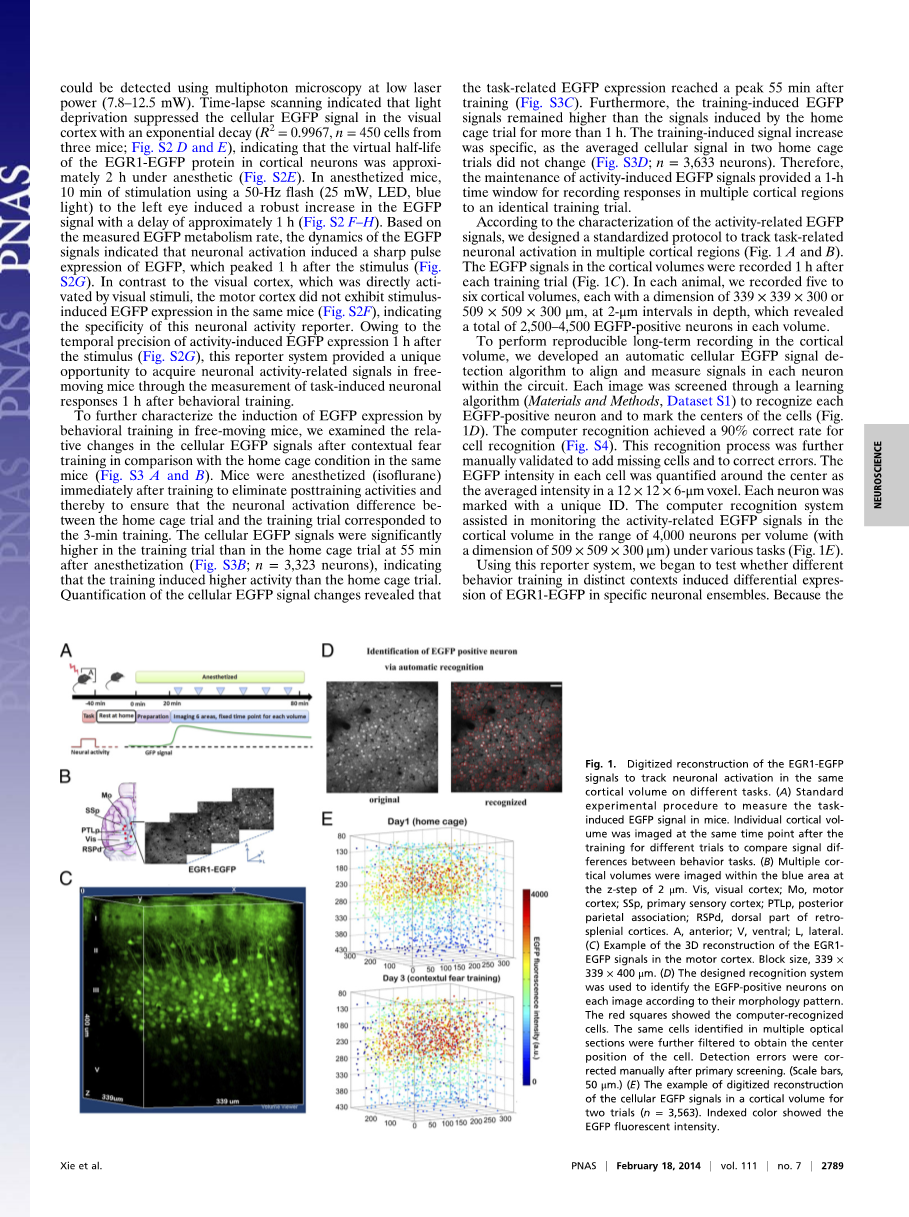
接下来,我们检测了小鼠受到视觉刺激时视皮层细胞EGFP信号的动态变化,EGFP信号通过一个颅窗采用多光子成像获取[30-32](Fig S2C)。在这一报告基因的鼠系中,EGR1-EGFP可以使用多光子显微镜以低激光功率检测(7.8-12。5mW)。延时扫描表明除去光会抑制视皮层细胞中的EGFP信号,使其呈指数式衰减(Rsup2;=0.9967,n==450,细胞来自三只小鼠;Fig S2Damp;E),表明在麻醉状态下皮层神经元中EGR1-EGFP蛋白的半衰期约为2小时(Fig S2E)。给予麻醉的小鼠左眼10分钟、50Hz的闪光刺激(25mW,LED,蓝光),会诱导其大约1小时后EGFP信号迅速增强(Fig S2F-H)。基于EGFP代谢速率的测量,EGFP信号的动态变化表明神经元的兴奋诱导EGFP急剧地表达,并在刺激后月1h达到峰值(Fig S2G)。与视皮层相比,同一小鼠在受到刺激后运动皮层并没有表现出刺激诱导的EGFP表达(Fig S2F),表明神经元活动报告基因的特异性。由于神经活动导致的EGFP表达信号精确的出现在刺激一小时后(Fig S2G),这一报告基因系统为自由活动的小鼠中获取神经活动信号提供了机会,可在行为训练一小时后测量任务诱导的神经元活动。
为了进一步研究自由活动的小鼠中行为训练诱导EGFP表达的特点,我们测试并对比了同一小鼠在条件恐惧训练后与在培养箱中细胞EGFP信号的相关变化(Fig S3Aamp;B)。在训练后,小鼠立即被麻醉(异氟烷)以消除训练后的神经活动,确保培养箱实验和训练实验间中神经元活动的差异都统一为三分钟训练。在麻醉55min后,训练实验组细胞EGFP信号显著高于培养箱实验(Fig 3B;n=3323个神经元),表明训练导致神经元活动较培养箱实验更高。细胞EGFP信号量的变化表明,训练相关的EGFP表达在训练后55min达到峰值(Fig S3C)。另外,训练诱导的EGFP信号在超过一小时的时间里持续高于培养箱实验。训练诱导的信号增强是特异性的,因为在两个培养箱实验中,平均细胞信号强度病么诶有改变(Fig S3D;n=3633个神经元)。因此,活动诱导的EGFP信号的持续存在为在多个皮层区域中记录神经元对同一训练实验的响应提供了一个小时的观测期。
根据活动相关的EGFP信号的特征,我们设计了一个标准的方案来追踪多个皮层区域汇中任务相关的神经元活动(Fig 1Aamp;B)。皮层卷中的EGFP信号在每次训练试验一小时后记录(Fig 1C),在每只小鼠中,我们记录了五到六个皮层卷,每个体积为339times;339
剩余内容已隐藏,支付完成后下载完整资料
资料编号:[235543],资料为PDF文档或Word文档,PDF文档可免费转换为Word
您可能感兴趣的文章
- 纳米材料介导镉污染土壤原位修复的机理及研究进展外文翻译资料
- 纳米零价铁磁洗受镉污染水稻土壤的有效性验证外文翻译资料
- 镉对水稻叶片生长和养分运输的短期影响外文翻译资料
- 间接竞争ELISA的研制及可视化基于金纳米棒生长的多色酶联免疫吸附试验玉米赤霉烯酮的测定外文翻译资料
- 铅(Pb)诱导的小麦根系生化和超微结构变化外文翻译资料
- 用蚕豆根尖微核试验评估污水灌溉区和生物修复区土壤的遗传毒性外文翻译资料
- 镧在红土壤中的生物累积和其对玉米幼苗生长的影响外文翻译资料
- 铈对水稻幼苗生长及部分抗氧化代谢的影响外文翻译资料
- 用黄体孢子菌沉淀碳酸盐固定有毒金属的体外研究及其在含硫化物尾矿中的应用外文翻译资料
- 水牛妊娠早期生物标志物的编码序列克隆及部分序列密码优化后在大肠杆菌中的表达外文翻译资料

