Sekiguchi Lesion Gene Encodes a Cytochrome P450 Monooxygenase That Catalyzes Conversion of Tryptamine to Serotonin in Rice
水稻编码细胞色素P450单加氧酶的SL基因在水稻中催化色胺转变为五羟色胺的研究
Tadashi Fujiwara , Sylvie Maisonneuve , Masayuki Isshiki , Masaharu Mizutani , Letian Chen ,Hann Ling Wong , Tsutomu Kawasaki , and Ko Shimamoto
(Receivedforpublication,December4,2009,andinrevisedform,February2,2010 Published,JBCPapersinPress,February11,2010,DOI10.1074/jbc.M109.091371)
前言:
5-羟色胺作为哺乳动物中常见的神经递质,在人类神经功能方面具有重要作用。在植物中,5-羟色胺的生物合成途径及其功能作用还不清楚。水稻sekiguchi lesion (sl)突变体积累色胺,而该物质是5-羟色胺生物合成的底物之一。通过图位克隆的方法,我们分离到一个SL基因,它编码细胞色素P450单加氧酶家族的CYP71P1蛋白。重组SL蛋白表现为5-羟化酶活性,催化色胺转变为5-羟色胺。该途径在哺乳动物未见报道,是一条新的5-羟色胺合成途径。研究表明,SL基因受到几丁质诱导物和水稻稻瘟病的诱导表达。外源施加5-羟色胺能诱导水稻悬浮组织防御基因表达和细胞死亡,且能提高植株对稻瘟病的抗性。此外,研究还发现RacGTPase和异源三聚体G 蛋白的Galpha;亚基介导了5-羟色胺诱导的防御反应。这些结果表明,5-羟色胺在水稻的先天免疫中起着重要作用。
正文:
植物发展了多种多样的免疫反应来应对病原菌,其中一种叫做超敏反应(HR),HR通常伴随细胞程序性死亡,它能高效抑制活体病原体进一步生长。HR通常伴随着活性氧的生成、细胞木质化、植物抗毒素生成和防御相关基因的表达。
为阐明HR在免疫反应中的作用,许多植物如水稻、拟南芥和玉米类病斑突变体被大量研究。类病斑突变体是一类在没有病原菌侵染下,自发性形成细胞死亡的一类突变体。水稻类病斑突变体sl病斑颜色橘黄色,病斑形成受到病原物包括稻瘟病的诱导,且病斑形成与色胺脱羧酶和单胺氧化酶活性有关。然而,SL基因的鉴定和功能研究及其在细胞死亡和植物抗病中的作用有待研究。
植物Rac/Rop GTPases在小GTPases的Rho家族中组成了一个特别的亚家族,该族蛋白在植物中高度保守。水稻中OsRac1调节一系列免疫反应,包括细胞死亡,活性氧生成,防御相关基因激活,细胞木质化和植物抗毒素生成。近来,发现OsRac1与HSP90、 HSP70、 OsMAPK6、 OsrbohB、Sti1/Hop和 RACK1形成的复合物在水稻先天免疫中发挥重要作用。异源三聚体G蛋白同样通过OsRac1在水稻先天免疫中发挥作用。事实上,水稻矮杆突变体d1由于缺少异源三聚体G 蛋白的Galpha;亚基,在防御反应诱导物刺激下其防御相关基因表达下降,且其更易感染稻瘟病。
5-羟色胺作为哺乳动物中重要的神经递质,参与了人类情绪、睡眠和焦虑的调节。尽管在许多植物中发现了5-羟色胺,但其功能未知。近来,报道称5-羟色胺参与水稻衰老和防御反应。此外,胡麻斑病和稻瘟病病斑部位的细胞壁中发现了5-羟色胺,表明5-羟色胺通过控制细胞壁的强度来限制病原菌的进一步侵染。
为了解SL基因在水稻细胞死亡和抗病性方面的作用,我们通过图位克隆分离到一个SL基因,该基因编码细胞色素P450单加氧酶,该酶催化色胺转变为5-羟色胺。此外,我们发现5-羟色胺通过激活细胞内的免疫反应信号途径来调控水稻先天免疫。
实验步骤
植物材料和基因定位——野生型粳稻品种Kinmaze和四种SL突变体(CM265,CM998,CM1019和CM2229),四种突变体为Kinmaze通过N-甲基-N-亚硝基脲诱变而来,以上SL突变体由日本国家水稻生物资源中心提供。CM265突变体与片段代换系SL263交叉,该片段代换系为粳稻Koshihikari背景,第12染色体带有籼稻Kasalath片段。利用STS和CAPS标记对F2和F3植株构建包含SL基因的物理图谱(见补充图S1)。详细定位信息见附加材料中实验步骤。
质粒构建和水稻转化——SL基因cDNA构建到具有玉米Ubp1启动子的p2K1载体上。为构建Sti1a/Hop RNA干扰载体,通过对Sti1a cDNA进行PCR扩增。并将其反向构建到具有Ubp1启动子的的PANDA载体上。OsRac1载体构建见相关文献。质粒构建细节见附件中的实验步骤。通过根癌农杆菌侵染水稻愈伤组织诱导转基因。
反转录和RT-PCR——利用TRlzol试剂(Invitrogen公司)来提取叶片和愈伤组织总RNA,RNA经DNaseI 处理,利用SuperScriptII反转录酶来合成cDNA,再用基因特异性引物来进行RT-PCR(见补充图S1)。
稻瘟病接种——在水稻4-6叶期喷雾浓度为105个孢子/ml的稻瘟病菌液,稻瘟病种为003和007。接种的幼苗在培养箱中24℃高温避光20h,然后转移至温室中,在特定的时间点取接种后的水稻叶片进行RT-PCR。为检验5-羟色胺对水稻稻瘟病抗性的影响,对水稻幼苗喷雾浓度为400ug/ml的5-羟色胺,22h按上述方法接种稻瘟病种007。接种6天后测量病斑长度。
几丁质、5-羟色胺、色胺处理——用2ug/ml几丁质诱导剂,400ug/ml(1.88mM) 5-羟色胺(5-羟色胺酸盐(Mr为212.68))或者400ug/ml(2.03mM)色胺(色胺酸盐(Mr为196.68))处理水稻诱导细胞一段时间。如前所述,少于30%的5-羟色胺用于培养实验。提取总RNA并用于RT-PCR。为检测细胞死亡,用5-羟色胺或色胺处理水稻悬浮细胞24h,用0.05%的伊文思蓝染色10min,然后用蒸馏水洗涤4次。用50%甲醇和1%SDS溶液在50℃中浸泡30min,死亡细胞会发生溶解,最后在595nm处测定吸光度值。
SL活性测定——SL和SL-G455D(CM265)cDNA克隆到pFastBac1载体(Invitrogen公司),然后转化到DH10Bac感受态大肠杆菌(Invitrogen公司)中。载体构建和转染等见产品手册。细胞Sf9异源表达和P450光谱测量参照前人方法。
SL或SL-G455D昆虫细胞微粒体表达载体构建方法如下:用磷酸缓冲液洗涤被感染的悬浮细胞(300ml),然后用buffer A(20mM的磷酸钾(pH值7.25),20%的(V/V)甘油,1mM EDTA和1mM二硫苏糖醇)重悬。超声处理细胞,10000times;g离心15min,除去细胞碎片。将上清液继续10000times;g离心1h,弃上清,用buffer A重悬,微粒体-80℃储藏用于进行酶活分析。
通过混合微粒体和拟南芥NADPH-P450还原酶来再生SL活性。反应体系包含20mM磷酸钾(pH 7.25),50pmol/ml重组P450蛋白,0.1unit/ml的NADPH- P450还原酶,1mM NADPH,和100uM色胺。反应通过加入NADPH来引发,在30℃下反应30min。样品用20mM磷酸盐(pH 2.5)稀释,然后HPLC4分析:柱子为COSMOSIL 5C18-PAQ柱(150mm长times;4.6mm内径),流速为1.0ml/min,在具有285nm的激发波长和在340nm的发射波长的荧光检测器中检测。5-羟色胺和色胺保留时间分别为6.5min和14.5min。用浓度为1~40uM的色胺底物来对SL进行动力学分析(通过加入NADPH来引起反应,在30℃下进行10min)。计算Km值和Kcat以及动力学常数。
免疫印迹——表达SL或SL-G455D蛋白的微粒体用附加材料实验步骤中的方法来做免疫印迹。
结果
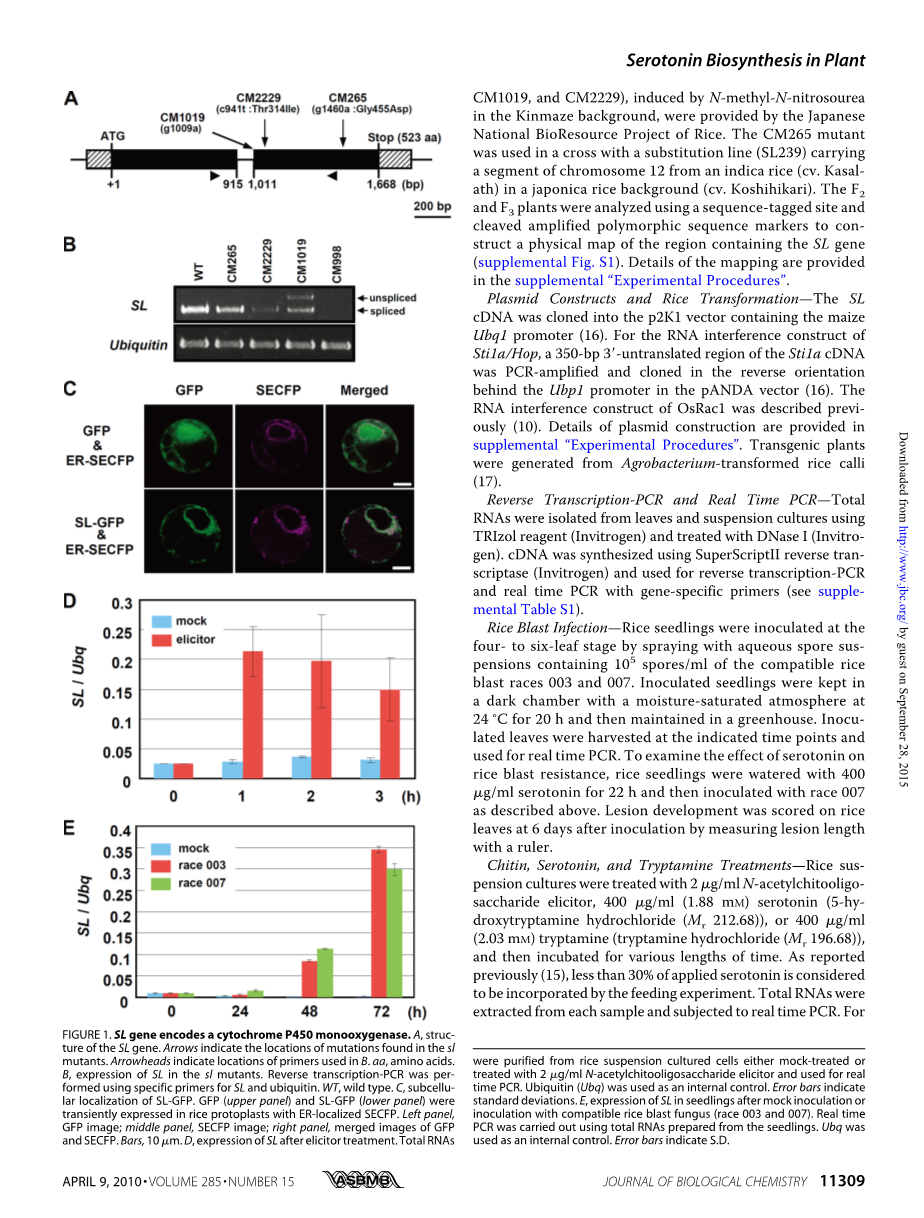
SL基因分离和图位克隆——为分离SL基因,我们通过N-甲基-N亚硝脲诱变获得了4个sl 突变体(CM265,CM2229,CM998和CM1019,见补充图S1)。sl 被定位到了第12号染色体350-kb范围内,里面含有7个候选基因(参考图S1)。PCR实验表明,CM998含有30-kb的缺失,这包含了一个P450蛋白和一个锌指结构蛋白的基因。对其他突变体测序,发现SL编码P450单加氧酶(图1A)。在CM2229中SL基因表达水平降低,由于CM998因为缺失整个P450基因,因此SL基因无表达。CM1019有两个可变转录本:3lsquo;端内含子的点突变导致了未剪切的转录本,另一个剪切靠近3rsquo;剪切位点,过表达P450 mRNA互补了SL表型(补充图S2)。
SL属于CYP71家族——SL蛋白存在血红素结合域, EXXR 序列, 以及富含脯氨酸的保守结构域(补充图S3)。CM265突变发生在血红素结合域的G455D突变位点,因为改变了其结构。CM2229第314位苏氨酸残基发生突变,而这一位点在P450循环中对氧化起着重要作用(补充图S3)。SL蛋白与鳄梨果CYP71A1蛋白有41%的相似性。在水稻CYP71家族中包含90个基因和28个假基因,然而,SL蛋白与同家族的任意蛋白相似性低于45%。SL蛋白是CYP71家族内的一类新蛋白,命名为CYP71P1。
利用水稻原生质体内的GFP-融合蛋白(图1C)的瞬时表达,定位SL蛋白在胞内的位置。SL蛋白被定位于内质网,在标记蛋白的N端添加了一个信号肽。结果表明,该信号肽被定位于内质网。
SL基因在先天免疫反应中的表达——几丁质诱导物(图1D)诱导水稻的防御方应,分析在水稻悬浮细胞中SL基因的表达。用几丁质诱导处理1h后可观察到SL mRNA水平明显上升。同样,接种稻瘟病菌后检测SL基因的表达水平。接种2种稻瘟病菌48h后,检测到SL基因表达,72h后SL基因表达进一步上升(图1E)。这些结果表明,SL在水稻先天免疫反应中发挥着作用。
SL蛋白具有色胺5-羟化酶活性 ——在以前的报道中称,SL突变体色胺含量比野生型高,而5-羟色胺含量很低。这说明SL催化色胺转变为5-羟色胺。为验证这一结论,构建了SL蛋白和SL-G445D蛋白的昆虫表达体系。通过免疫印迹检验重组SL蛋白和SL-G445D蛋白的表达(图2A)。包含野生型SL蛋白的片段在450nm处有明显的吸收峰,而包含SL-G445D蛋白的片段无吸收峰出现(图2B)。这些结果说明,SL蛋白是细胞色素P450酶。SL重组蛋白进一步用于酶活性分析。HPCL分析发现,表达SL蛋白的昆虫表达体系中检测到了5-羟色胺,而SL-G445D中未检测到。SL蛋白高效专一催化色胺5rsquo;羟基化(Km =
7.3 0.8 uM and kcat =45 1.5 min-1),SL蛋白质没有代谢产生色氨酸,吲哚-3-乙酸,或酪胺。这说明SL蛋白是色胺5-羟基化酶,它年能催化色胺转变为5-羟色胺。
5-羟色胺在先天免疫反应中具有重要作用——在以前的报道中,在水稻胡麻斑病和稻瘟病病斑处有大量5-羟色胺积累。而在sl突变体中却没有5-羟色胺生成。此外,sl突变体比野生型更易感染胡麻斑病,施加5-羟色胺后,sl突变体病斑病位的病原菌生长受到抑制。这说明5-羟色胺可能参与先天性免疫。为阐明5-羟色胺如何调控先天免疫,我们对4个防御基因PBZ1、PAL1、Cht1、Cht3在水稻悬浮组织中的表达进行了分析,
剩余内容已隐藏,支付完成后下载完整资料

英语原文共 7 页,剩余内容已隐藏,支付完成后下载完整资料
资料编号:[286436],资料为PDF文档或Word文档,PDF文档可免费转换为Word
您可能感兴趣的文章
- 纳米材料介导镉污染土壤原位修复的机理及研究进展外文翻译资料
- 纳米零价铁磁洗受镉污染水稻土壤的有效性验证外文翻译资料
- 镉对水稻叶片生长和养分运输的短期影响外文翻译资料
- 间接竞争ELISA的研制及可视化基于金纳米棒生长的多色酶联免疫吸附试验玉米赤霉烯酮的测定外文翻译资料
- 铅(Pb)诱导的小麦根系生化和超微结构变化外文翻译资料
- 用蚕豆根尖微核试验评估污水灌溉区和生物修复区土壤的遗传毒性外文翻译资料
- 镧在红土壤中的生物累积和其对玉米幼苗生长的影响外文翻译资料
- 铈对水稻幼苗生长及部分抗氧化代谢的影响外文翻译资料
- 用黄体孢子菌沉淀碳酸盐固定有毒金属的体外研究及其在含硫化物尾矿中的应用外文翻译资料
- 水牛妊娠早期生物标志物的编码序列克隆及部分序列密码优化后在大肠杆菌中的表达外文翻译资料

