JAV1调控茉莉酸介导的植物抗性反应
Po Hu,1 Wu Zhou,1 Zhiwei Cheng,1,2 Meng Fan,1 Lei Wang,3 and Daoxin Xie1,*
1Tsinghua-Peking Center for Life Sciences, MOE Key Laboratory of Bioinformatics, School of Life Sciences, Tsinghua University, Beijing 100084, China
2Key Laboratory of Integrated Pest Management in Crops, Institute of Plant Protection
3Biotechnology Research Institute
Chinese Academy of Agricultural Sciences, Beijing 100081, China
*Correspondence: daoxinlab@tsinghua.edu.cn
http://dx.doi.org/10.1016/j.molcel.2013.04.027
摘要
植物进化出有效的机制来保护自己免受环境胁迫,并使用茉莉酸作为防御昆虫攻击和病原体感染的重要防御信号。茉莉酸也被认为是一种重要的生长调节因子,通过它可以调节多种发育过程。尽管进行了大量研究,但尚未报道茉莉酸调节的独立于发育反应的植物防御关键信号成分。我们鉴定了JAV1,它是茉莉酸途径中的关键基因,其作为控制植物防御的负调节因子起作用,但在植物发育中不起作用。我们的研究结果表明,当遇到昆虫攻击和病原体感染时,植物会积累茉莉酸酯,通过26S蛋白酶体引发JAV1降解,从而激活防御基因表达并提高对昆虫和病原体的抗性。这些发现提供了植物积累茉莉酮酸信号以保护自身免受昆虫攻击和病原体感染的分子机制。
引言
高等植物在其生命周期中经常遇到环境胁迫,例如昆虫和病原体的攻击。除了组成性的物理和化学策略,植物已经开发出强大的诱导防御系统来抵御多种攻击者(Glazebrook,2005; Howe和Jander,2008;Jones和Dangl,2006)。植物防御系统在很大程度上取决于大量防御基因的诱导表达。许多前线防御反应基因的表达受茉莉酮酸酯的控制(Creelman和Mullet,1997; Reymond等,2000),茉莉酸是一类环状脂肪酸衍生的植物激素,在结构上类似于动物防御调节剂前列腺素(Creelman) 和Mullet,1997)。
茉莉酸,包括茉莉酸及其环戊酮衍生物(Creelman和Mullet,1997; Farmer等,2003),已成为调节植物对食草昆虫和坏死性病原体的反应的重要防御信号分子。它们还是一种重要的生长调节剂,可以调节植物发育过程的各个方面,如植物育性、块茎化和果实成熟(Browse,2009; Cheong and Choi,2003; Creelman和Mullet,1997; Erb等,2012; Farmer等,2003; Liechti和Farmer,2002; Wasternack和Kombrink,2010)。为了响应发育信号和环境压力,茉莉酮酸酯在植物中被有效地合成和积累(Li et al。,2002; Waste-rnack and Kombrink,2010)。冠毒素不敏感1(COI1)(Xie等,1998)感知茉莉酮酸信号(Sheard等,2010; Yan等,2009)并募集茉莉酮酸酯-ZIM结构域蛋白(JAZs)(Chini等 ,2007; Thines等,2007; Yan等,2007)通过26S蛋白酶体途径进行泛素化和降解,导致各种转录因子的激活,随后调节其下游信号级联响应并调节相应的植物反应。R2R3-MYB转录因子(MYB21和MYB24)和转录复合物WD-repeat / bHLH / MYB与JAZs相互作用,特异性调节茉莉酸介导的植物育性(Song et al。,2011,2013),色素沉着和毛状体发生( Qi et al。,2011),而基本螺旋 - 环 - 螺旋(bHLH)转录因子(MYC2,MYC3和MYC4)是重要的信号组分,在防御反应的调节(Dombrecht等, 2007; Ferna ndez-Calvo等,2011; Wild等,2012)和各种发育过程,如茉莉酮酸抑制根生长(Chen et al。,2011; Cheng et al。,2011; Ferna ndez-Calvo et al。,2011; Niu et al。,2011),根茎细胞生态位发育(Chen et al。,2011),倍半萜积累(Hong et al。,2012),和光致形成幼苗生长(Yadav等,2005)中具有双重作用。迄今为止报道尚未确定任何关键信号因子,以独立于发育调节的方式特异性控制茉莉酮酸介导的防御反应。因此,目前不可能通过操纵茉莉酮酸信号通路有效地改良植物对昆虫和病原体的抗性。
在这项研究中,我们构建了一个拟南芥转基因RNAi文库,并通过遗传筛选20,000个转基因RNAi文库鉴定了JAV1基因。沉默JAV1基因可显着增强茉莉酸调节的对抗营养性病原体和食草昆虫的防御反应,但不会严重改变茉莉酸酯介导的发育过程。我们的研究结果提供了一种分子机制观点,当昆虫和病原体攻击时,植物通过26S蛋白酶体积累茉莉酸酯诱导JAV1蛋白降解,随后增强各种免疫应答基因的诱导表达,从而激活植物免疫系统,以防御昆虫袭击和病原体感染。
结果
通过遗传筛选拟南芥转基因RNAi文库鉴定JAV1
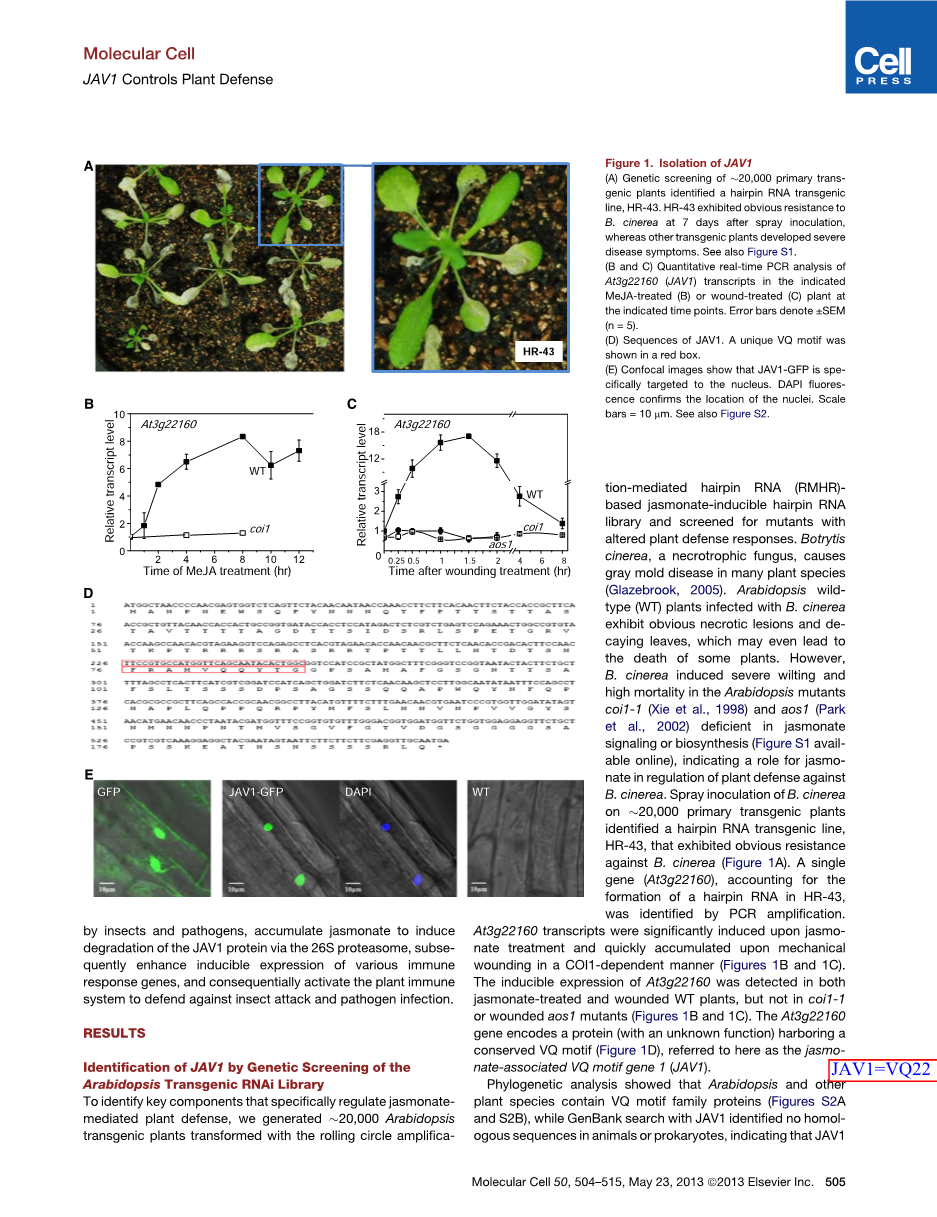
为了确定特异性调节茉莉酮酸介导的植物防御的关键组分,我们构建了20,000个用滚环扩增介导的基于发夹RNA(RMHR)的茉莉酸-诱导型发夹RNA文库转化转基因植物,并筛选具有改变的植物防御反应的突变体。灰葡萄孢(Botrytis cinerea)是一种死体营养型真菌,在许多植物物种中引起灰霉病(Glazebrook,2005)。用Bcinerea感染的拟南芥野生型(WT)植物表现出明显的坏死病灶和脱落叶,这甚至可能导致一些植物的死亡。然而,灰霉病引起突变体coi1-1(Xie等人,1998)和aos1(Park等人,2002)叶卷曲及高死亡率,这些突变体在茉莉酮酸信号传导或生物合成方面存在严重缺陷、(图S1可在线获得),表明茉莉酸在植物防御B.cinerea中的作用。通过PCR扩增鉴定了在HR-43中形成发夹RNA的单个基因(At3g22160)。在茉莉酸盐处理时显着诱导At3g22160转录表达并且在机械伤害时以COI1依赖性方式快速累积(图1B和1C)。在茉莉酮酸处理的和受伤的WT植物中均检测到At3g22160的诱导型表达,但在coi1-1或受伤的aos1突变体中未检测到(图1B和1C)。At3g22160基因编码具有保守VQ基序的蛋白质(具有未知功能)(图1D),在此称为与茉莉酸相关的VQ基序1(JAV1)。
图1.JAV1的隔离
(A)对20,000个初级转基因植物进行遗传筛选鉴定了发夹RNA转基因株系HR-43。HR-43对灰霉菌表现出明显的抗性。HR-43在喷雾接种后7天表现出对灰霉菌的明显抗性,而其他转基因植物则出现严重的感病症状。另见图S1。
(B和C)在指定的MeJA处理的(B)或伤口处理的At3g22160(JAV1)转录物的定量实时PCR分析
(C)植物中对At3g22160(JAV1)转录物进行定量实时PCR分析。误差棒表示plusmn;SEM(n = 5)。
(D)JAV1的序列。一个独特的VQ结构域显示在一个红色框中。
(E)共聚焦图像显示JAV1-GFP特异性靶向细胞核。DAPI荧光证实了细胞核的位置。比例尺=10毫米。另见图S2。
系统发育分析表明,拟南芥和其他植物物种含有VQ基序家族蛋白(图S2A和S2B),而用JAV1进行的GenBank搜索在动物或原核生物中未发现同源序列,表明JAV1可能是植物特异性的。对推定的JAV1序列的分析突出了潜在的核定位信号(NLS),包括SV40大T抗原和O2MT,分别对应于单分子和二分子核靶向信号(图S2C)。检查表达GFP融合的JAV1的转基因拟南芥的亚细胞定位表明它是核定位的(图1E)。
图2. JAV1 RNAi转基因系的特性
(A)在转基因品系Ri17和Ri55中,JAV1基因的转录水平显着降低,而其他含VQ结构域的基因未受影响。通过定量实时PCR确定相对转录物水平。三个含有VQ基序的基因(At1g80450,At2g41010和At4g15120)的数据在图S2A中用黑色星标记显示。误差棒表示plusmn;SEM(n=5)。
(B)JAV1基因的转录水平在另外8个转基因JAV1 RNAi系中显着降低。误差棒表示plusmn;SEM(n=5)。
(C)JAV1不参与茉莉酮酸介导的植物根生长。来自WT和JAV1 RNAi转基因系Ri17和Ri55的幼苗在没有(上图)或具有(下图)25mM MeJA的MS平板上生长7天。比例尺=5毫米。
(D)(C)中所示植物的相对根长。误差棒表示plusmn;SEM(n=3)。
(E)代表性地显示了WT和Ri17的花,花序和种子的表型。
JAV1基因控制茉莉酸介导的植物防御对病原体感染的反应
为了证实HR-43中疾病抗性的增加是发夹RNA沉默JAV1的结果,我们生成了JAV1RNAi转基因株系,其显示JAV1转录水平特异性降低但不含其他含VQ基序的基因(图2A和2B)。JAV1RNAi转基因品系Ri17和Ri55分别保持10%和20%的野生型JAV1水平(图2A)。在Ri17,Ri55或其他独立的JAV1 RNAi转基因株系中,形态学表型,包括根生长、砧孔、花、种子和其他发育的过程,没有明显受到影响(图2C-2E和3A以及数据未显示),表明JAV1在茉莉酮酸调节的植物生长和发育中不起重要作用。
为了检查在JAV1 RNAi转基因品系中对B.cinerea的植物防御反应,用B.cinerea孢子悬浮液(2.53times;104孢子/ml;2ml/植物)喷洒WT,coi1,Ri17和Ri55并在一致条件下生长。潮湿或潮湿的条件下7天。通过表型观察,相对鲜重,病情严重程度或病变面积以及植物存活百分比来测量疾病症状。在全植物喷雾接种试验中,Ri17和Ri55对灰霉病菌表现出明显的抗性并且疾病症状较少,而coi1突变体植物对白粉虱感染更敏感,相对鲜重显示(coi1为32%,WT为68%,Ri17和Ri55均为88%),疾病严重程度(coi1为85%,WT为42%,Ri17和Ri55为20%),植物存活率(coi1为42%,80%)对于WT,以及对于Ri17和Ri550都是100%(图3A-3D)。用单独的叶子上的5ml灰葡萄孢悬浮液(2.5times;10 4孢子/ml)进行接种实验;在coi1和WT中发展出明显的疾病症状,但在Ri17和Ri55品系中没有(图3E和3F)。这些结果表明JAV1基因调节茉莉酸酯介导的植物对抗坏死真菌B.cinerea的抗性。
JAV1控制茉莉酸-调节植物防御昆虫攻击
为了研究JAV1是否参与茉莉酮酸调节的植物防御昆虫攻击,将WT,coi1-1和JAV1RNAi转基因品系的成熟莲座叶饲喂甜菜夜蛾(Spodoptera exigua),这是一种具有广泛宿主范围的全球重要农业害虫(Howe和Jander,2008年)。当喂食coi1植株时,甜菜幼虫吞噬了大部分叶子(图4A),生长并迅速发育(图4B和4C)。相比之下,Ri17和Ri55显示甜菜夜蛾的消耗减少(图4A)并且缩减了甜菜夜蛾幼虫的生长(图4B和4C)。当甜菜夜蛾幼虫用8个另外独立产生的JAV1RNAi转基因品系饲喂时获得相同的结果(图S3A和S3B)。类似地,Ri55和Ri17表现出对抗Bradysia impatiens的防御,这是一种主要以腐烂的植物物质为食的普遍害虫。大多数coi1植物在感染后20天死于B.impatiens幼虫攻击;WT植物也被B.impatiens严重侵染。然而,Ri17和Ri55中的植物存活率显着更高(图4D和S3C),表明沉默JAV1提高植物对B.impatiens攻击的防御。
蚜虫(Myzus persicae Su ̈ lzer)是破坏许多重要作物的主要害虫(Howe和Jander,2008)。在双选择测试中,JAV1 RNAi系Ri17容纳较少的蚜虫,而当蚜虫被选择在Ri17和coi1或Ri17和WT植物之间进行选择时,coi1和WT植物吸引更多的昆虫(图4E)。在Ri17和coi1-1之间蚜虫的分布频率为25%和75%,在Ri17和WT之间为42%和58%(图4E)。在非选择性试验中,在WT,coi1和Ri17的每株植物上释放20只蚜虫。分别接种48小时后计算每株植物上蚜虫(成虫 若虫)的数量,表明Ri17植物显示出对蚜虫发育的明显抑制(图4F)。总之,这些结果表明JAV1调节植物对抗蚜虫的防御。
图3.JAV1控制茉莉酸介导的植物对B.cinerea的防御
(A)用灰霉菌或水(CK)喷雾接种后7天,WT,coi1-1和两种JAV1RNAi转基因植物(Ri17和Ri55)的表型。 剩余内容已隐藏,支付完成后下载完整资料

英语原文共 12 页,剩余内容已隐藏,支付完成后下载完整资料
资料编号:[278939],资料为PDF文档或Word文档,PDF文档可免费转换为Word
您可能感兴趣的文章
- 纳米材料介导镉污染土壤原位修复的机理及研究进展外文翻译资料
- 纳米零价铁磁洗受镉污染水稻土壤的有效性验证外文翻译资料
- 镉对水稻叶片生长和养分运输的短期影响外文翻译资料
- 间接竞争ELISA的研制及可视化基于金纳米棒生长的多色酶联免疫吸附试验玉米赤霉烯酮的测定外文翻译资料
- 铅(Pb)诱导的小麦根系生化和超微结构变化外文翻译资料
- 用蚕豆根尖微核试验评估污水灌溉区和生物修复区土壤的遗传毒性外文翻译资料
- 镧在红土壤中的生物累积和其对玉米幼苗生长的影响外文翻译资料
- 铈对水稻幼苗生长及部分抗氧化代谢的影响外文翻译资料
- 用黄体孢子菌沉淀碳酸盐固定有毒金属的体外研究及其在含硫化物尾矿中的应用外文翻译资料
- 水牛妊娠早期生物标志物的编码序列克隆及部分序列密码优化后在大肠杆菌中的表达外文翻译资料

