
英语原文共 15 页,剩余内容已隐藏,支付完成后下载完整资料
基于“原子”层面的方法对强化生物除磷微生物学研究的贡献
Christina M. Forbes1, Niall D. Orsquo;Leary1 , Alan D. Dobson1 amp; Juian R. Marchesi2
1Department of Microbiology, Environmental Research Institute, University College Cork, Cork, Ireland; and 2School of Biosciences, Cardiff University, Cardiff, UK摘要:自30多年前首次进行研究以来,微生物在废水中生物去除磷酸盐方面的作用一直受到关注。基于“原子”理论的最新进展极大地增进了我们在这一领域的认知,并促进了对现有强化生物除磷( EBPR )模型的改进,这些模型主要以研究该过程的独立培养为基础。基于这种方法对EBPR的理解 ,这一精妙观点将集中表现在我们对EBPR过程的最新进展上。
关键词:强化生物除磷;“原子”方法;独立培养
引言
近年来,越来越严格的环境立法加大了对废水中无机营养物水平的控制,如磷和氮,这些无机营养物可以在排放许可标准下排放到自然水体中。这些营养元素在家庭废水、农业径流和一些食品加工废水中含量很高(Danalewich et al., 1998; McGrath et al., 2001)。长期以来,人们一直认为这些营养物质含量的提高对淡水环境的质量有害。尤其是磷,被认为是一个主要因素,如今,人们已经意识到磷是大多数湖泊和河流中真核藻类生长的主要限制因素( Schindler,1977年)。目前,化学和生物方法都被广泛用于磷酸盐的去除。然而,越来越多的证据表明使用生物处理有效去除磷,是一种成本效益高、环境可持续的方法,可以将污水磷浓度降低到符合欧洲标准的限度内。欧洲理事会第91 / 271号城市废水处理指令和第98 / 15号修正案概述了这些限制,其中规定,排放到敏感淡水区域的废水必须经过处理,以使磷酸盐减少80 %,或者最终磷酸盐浓度在1 - 2毫克每升之间。根据淡水水体对富营养化的敏感性,以及其制造饮用水的潜在用途,淡水水体被称为“敏感性”水体。
强化生物除磷
废物流中的生物除磷是通过利用一些生物在超过正常代谢需求的情况下生物积累胞内多聚磷酸盐的能力来实现的。这种现象最初被称为“奢侈”磷吸收(Levin amp; Shapiro, 1965),但现在这一过程通常被称为强化生物除磷。强化生物除磷( EBPR )用于多种废物流的修复已经超过30年,通常在序批式反应器( SBR )中进行,该反应器在厌氧和好氧阶段之间循环活性污泥(Barnard, 1974; Mino amp; Satoh, 2006)。这种“双相”方法有选择地产生聚磷酸盐积累生物体( PAOs ),这些生物体能够适应EBPR过程中磷酸盐的“富足和稀缺”的性质,同时伴随着生物累积磷酸盐。在该过程的厌氧阶段,富含磷酸盐的废水被应用于活性污泥,生物质中的PAOs快速吸收碳底物,最常见的挥发性脂肪酸( VFAs ),形成聚- B-羟基烷酸酯( PHA ) ( Daws amp; Advanced,1973年;图1 )。在来自实验室规模程度的研究中,乙酸盐被广泛用作EBPR的碳源;已经证明PHA的化学成分取决于被同化的VFA底物(Hesselmann et al., 1999; Levantesi et al., 2002; McMahon et al., 2002)。这些聚合物的合成伴随着细胞内多聚磷酸盐磷酸酐键的断裂、ATP的生成以及金属螯合正磷酸盐从细胞中的释放(Kornberg, 1995; Kulaev amp; Kulakovskaya, 2000)。现在普遍认为,产生还原聚合物PHA所需的能量来源于胞内糖原的降解,而相应的NADH则通过Embden - Meyerhof ( EM )或Entner - Doudoroff ( ED )途径产生。然而,单独糖原降解不能为以乙酸盐为碳源的EBPR产生足够的还原潜力,三羧酸( TCA )循环被认为产生了所需的额外还原能力( Schuler amp; Jenkins,2003; Seviour et al., 2003)。
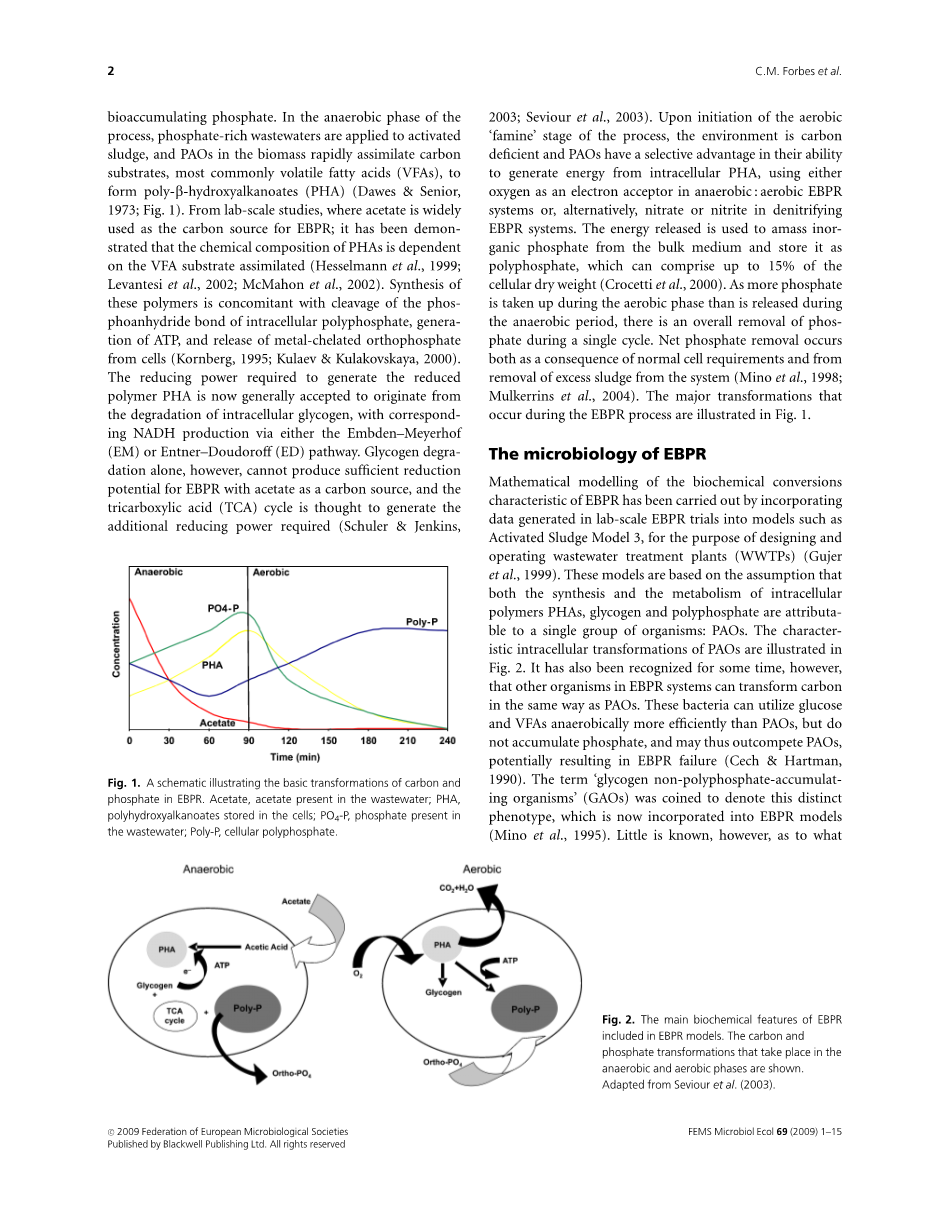
图1。EBPR中碳和磷酸盐基本转化的示意图。乙酸盐,废水中存在的乙酸盐;PHA,储存在细胞中的聚羟基链烷酸酯; PO4-P,废水中存在的磷酸盐;聚磷,多孔聚磷酸盐。
在该过程的有氧“稀缺”阶段开始时,环境是缺碳的,PAO在利用 细胞内的PHA产生能量方面具有选择性优势,在厌氧环境下将氧作为电子受体,即好养EBPR系统,或者在反硝化系统中将硝酸盐或亚硝酸盐作为电子受体。释放的能量被用于从散装介质中积聚无机磷酸盐,并以多磷酸盐的形式储存,多磷酸盐可占细胞干重的15%(Crocettietal., 2000)。由于需氧阶段吸收的磷酸盐比厌氧阶段释放的多,因此在单个循环中磷的去除是全面的。净磷酸盐去除既是正常细胞需求的结果,也是从系统中去除过量污泥的结果(Minoetal., 1998; Mulkerrins et al., 2004)。EBPR过程中发生的主要转变如图1所示。
EBPR的微生物学
EBPR生化转化特性的数学模型是通过将实验室规模EBPR试验中产生的数据纳入活性污泥模型3等模型中来进行的,目的是设计和运营污水处理厂( WWTPs ) (Gujer et al., 1999)。这些模型基于这样的假设,即细胞内聚合物PHA、糖原和多聚磷酸盐的合成和代谢都可以归因于一组生物: PAOs。PAOs的特征性细胞内转化如图2所示。然而,一段时间以来人们也认识到,EBPR系统中的其他生物可以像PAOs一样转化碳。这些细菌可以比PAOs更有效地厌氧利用葡萄糖和VFAs,但不会积累磷酸盐,因此可能会超过PAOs,可能导致EBPR失败(Cech amp; Hartman, 1990)术语“糖原非聚磷酸盐积累生物体”( GAOs )是为了表示这种独特的表型而发明的,这种表型现已被纳入EBPR模型( Mino等人,1995年)。
图2。EBPR的主要生化特征包括在EBPR模型中。显示了在厌氧和好氧阶段发生的碳和磷转化。改编自Seviour等人( 2003)。
然而,是什么实验条件导致了这些生物在PAOs上增殖,人们知之甚少。(Fukase et al., 1985; Cech amp; Hartman, 1990, 1993; Liu et al., 1994; Matsuo, 1994; Satoh et al., 1994; Sudiana et al., 1999)。缺乏对促进GAO增长因素的根本理解就意味着对EBPR过程的良好控制并不总是容易实现的,并且当问题确实发生时,补救过程失败的措施并不总是可靠的( Mino等人,1998年)。更好地理解EBPR的微生物学,平衡有利于PAO增殖的条件,因此EBPR过程的成功至关重要。
在过去的30年里,大量的努力都集中在识别在EBPR过程中起关键作用的生物,以及确定这些生物盛行的条件上。这方面最早的尝试涉及从EBPR污泥中直接分离聚氧乙烯。第一次成功培养显示出高磷酸盐去除能力的生物的尝试导致了球菌状细胞的分离,这些细胞被鉴定为Gammaproteo细菌不动杆菌属的成员( Fuhs amp; Chen,1975年)。从EBPR生物量中分离聚磷酸盐积累不动杆菌是容易实现的,并且得出结论,它们在EBPR生物量中数量很大。(Deinema et al., 1980, 1985; Buchan, 1983; Luml;otter, 1985; Wentzel et al., 1988; Beacham et al., 1992)。鉴于这些发现,EBPR中的不动杆菌属成员被鉴定为除磷细菌,并将其命名为“PAO”。然而,对这些不动杆菌分离株的生化研究表明,它们不具备PAOs的特征代谢属性(Tandoi et al., 1998),也没有足够数量的磷酸盐去除剂(Wagner et al., 1994)。使用直接培养技术,各种系统发育上不同的生物体随后被提出为PAOs。微小的磷细菌、蓝细菌和其他细菌被培养,并被证明能够在细胞内积累磷(Ubukata amp; Takii, 1994; Nakamura et al., 1995; Stante et al., 1997)。然而,当进一步调查时,这些分离物在EBPR中的作用就不可信了。举例来说,已经证明了蓝细菌的形态特征不同于PAO的形态特征,而磷在细胞内转化也与PAO不一致。
“原子”技术与EBPR
从一开始,“基于原子的技术”就使微生物生态学领域发生了革命。Staley amp; Konopka ( 1985 )的平板计数异常预示着复杂微生物环境研究的一个新阶段,随着PCR的出现,一个新的发现时代开始于独立培养技术的发展。在复杂微生物生态系统中识别生理相关生物的努力将重点转向了16S rRNA基因的表征,该基因已被确定为识别和量化生物多样性的合适目标,从而允许更详细的微生物生态系统特征( Woese amp; Fox,1977年)。独立于培养技术在EBPR研究领域的最早应用包括使用荧光标记rRNA靶向寡核苷酸探针原位探测EBPR污泥。Wagner等人 ( 1994年)开发了一种不动杆菌特异性探针,并将其应用于磷酸盐积累生物量,结论是该属仅占污泥细菌总数的10 %,因此,不仅仅是磷酸盐去除的原因。据指出,来自beta;蛋白细菌和高G C革兰氏阳性菌(放线菌)的细胞与聚磷酸盐内含物相关,因此很可能是EBPR中的聚磷酸盐。正在研究的EBPR污泥中分离物的生长表明不动杆菌属是最容易培养的菌株,占可培养细菌的32 - 33 %。这一发现显示了独立培养的研究的固有偏见,并强调了该领域独立培养方法的令人兴奋的潜力。从那时起,利用各种基于“原子”的方法,对参与EBPR的PAO的搜索进展迅速。基于基因组的“微生物学”技术已经被证明是特别有用的工具,既可以辨别活性污泥中被认为是PAOs的分离物的作用,也可以帮助识别在EBPR过程中具有生理相关性的新生物。据报道,通过将“微生物学”技术应用于EBPR生物量,多种生物在EBPR的成败中扮演着关键角色。
PAOs
在Wagner等人的研究之后。( 1994年),其他研究小组使用同样的策略来阐明EBPR污泥中的PAO。另外,通过利用FISH,Kampfer等人( 1996年)还得出结论,Betaproteobacteria和放线菌是EBPR生物量中的主导群体,与DAPI染色的多磷酸盐夹杂物有关。在进一步的研究中,其他基于16S rRNA基因的技术被用于研究EPBR污泥,在一项涉及EBPR污泥的16S rRNA基因克隆文库构建的研究中,放线菌再次被提议为主导(Christensson等人,1998年)。在一项利用末端限制性片段长度多态性分析研究EBPR生物量的研究之后,得出了同样的结论( Liu等人,1998年)。
Bond等人的一项研究支持了对Betaproteobacteria作为EBPR中对PAOs的见解。他们( 1995年)生成两个16S rRNA基因克隆文库进行分析除磷污泥和非除磷污泥中的细菌联合体,并得出结论,与红循环菌属相关的细菌最有可能是PAOs。在研究的两种污泥中,尽管beta;-变形菌最丰富,但在除磷生物量中,beta;-变形菌红循环群的频率较高。Bond等人随后报道了对EBPR有反应的beta;蛋白细菌系统发育的更具体信息。( 1999年)。在这项研究中,FISH被用来检测各种活性污泥的细菌群落结构,每种污泥都表现出不同程度的除磷性能。据指出,显示出良好磷酸盐去除的污泥主要由蛋白细菌的b-2亚类(占总细菌的55 % )和放线菌(占总细菌的35 % )控制。相比之下,除了b-1和b-2亚类之外,除了B- 1和B- 2亚类之外,表现出较差磷酸盐去除能力的污泥主要由beta;蛋白细菌控制。Hesselmann et al. ( 1999年)使用全周期RNA方法对类似红环菌的PAOs进行了更详细的分类。在本研究中,观察到在乙酸盐上培养的EBPR污泥主要由beta;蛋白细菌中与红环类相关的细胞系统发育所控制。通过克隆从这种生物量中扩增的16S rRNA基因,检索了两个主要的与红循环相关的序列。特定探针RHC 175和RHC 439被设计成靶向这些序列,并且在应用于生物质之后,注意到这些序列中的一个,名为lsquo;R6rsquo;,主导生物质。胞内多羟基丁酸酯和多聚磷酸盐的化学染色证实,这种类型的生物化学与EBPR代谢模型非常吻合。尽管与红环藻有密切关系,但这种被认为是PAO的植物并没有光养生长,这就排除了它作为红环藻属成员的分类。鉴于PAO不能被分离出来,因此不能完全表征,术语“念珠菌”被用来描述该生物以及临时的新属和新种“伏隔菌磷脂”(以下简称伏隔菌)。Crocetti等人( 2000年)随后使用结构域、分裂和亚区级特异性探针证实了这种物种在EBPR中的重要性。超过80 %的细菌被鉴定为含有多磷酸盐的b-2蛋白细菌,它们排列成球菌属。16S rRNA基因克隆文库由受检污泥构建,检索到的序列表明PAO可能是伏隔菌。名为PAO462、PAO651和PAO846的DNA探针被设计成针对该候选PAO,并随后被用于分析显示不同程度磷酸盐去除的大量污泥,这使得存在的探针结合细胞的数量和被研究生物质的磷酸盐去除能力之间可以得出关联。
Zilles(2002)等人随后证明了伏隔菌在整个系统中的重要性。在本研究中,FISH被用来利用序列特异性探针分析了三种典型全尺寸构型的污水处理厂的污泥。来自多种WWT植物的红环菌属的生物量是可观的,尽管已经证明EBPR过程的处理方式培养出了不同细菌群体,但发现与红环菌相关的生物有机体占污泥中存在的细菌群体的26 %至73 %。在随后的一些研究中,使用特定的探针进一步评估
全文共38027字,剩余内容已隐藏,支付完成后下载完整资料
资料编号:[801]

